Le vieillissement biologique
Plus qu'être un réel travail de recherche, ce qui suit reprend plutôt librement les idées que le professeur Steven Austad expose dans son passionnant livre [1], dont je conseille vivement la lecture à toute personne intéressée par le sujet.
Vieillir est encore le seul moyen qu'on ait
trouvé de vivre longtemps.
Charles Augustin Sainte Beuve
Tout le monde désire vivre longtemps, mais personne ne voudrait
être vieux.
Jonathan Swift
| Drosophile | 0,1 |
| Souris | 3 |
| Anguille | 6 |
| Dinde | 12,5 |
| Chèvre | 18 |
| Chauve-souris vampire | 19,5 |
| Chat | 21 |
| Canari | 24 |
| Salamandre tachetée | 25 |
| Lion | 29 |
| Boeuf | 30 |
| Grizzly | 31 |
| Petite chauve-souris brune | 32 |
| Chien | 34 |
| Pigeon | 35 |
| Chimpanzé | 37 |
| Esturgeon | 50 |
| Alligator | 56 |
| Éléphant | 57 |
| Cheval | 62 |
| Ara | 64 |
| Homme | 90 |
| Tortue géante | 177 |
Longévité maximale constatée (en
années) de quelques espèces animales
(Sources :
Encyclopædia Britannica
,
Quid et [1])
Vieillissement et longévité
Il s'agit dans un premier temps de définir précisément le vieillissement
biologique, que l'on appelle aussi la sénescence, et d'en donner
des indicateurs fiables. Steven N. Austad [1] donne
la définition suivante du vieillissement : la détérioration
progressive de la quasi-totalité des fonctions de l'organisme au cours du
temps
.
Le premier critère qui vient à l'esprit pour caractériser le vieillissement semble être la longévité ou espérance de vie, c'est-à-dire la durée de vie moyenne d'un individu. Cependant, il est clair que cette donnée ne dépend pas seulement du vieillissement, mais aussi de critères environnementaux. Considérons, comme le fit le prix Nobel de médecine Peter Medawar, l'hypothétique exemple, riche en enseignements, d'une population de tubes à essai dans un laboratoire. Si l'on néglige les fêlures qui peuvent éventuellement affecter leur fragilité, ces tubes ne « vieillissent » pas. Cependant, la pression environnementale représentée ici par les manipulations des chercheurs et laborantins conduira fatalement à la casse de chacun des tubes à essai. On peut donc tout à fait parler de la longévité d'un tube à essai, mais celle-ci n'a aucun lien avec un quelconque vieillissement intrinsèque mais dépend exclusivement de l'habileté des expérimentateurs. Ainsi, suivant les laboratoires, l'espérance de vie d'un tube à essai peut être multipliée par dix. Il en est de même dans les populations animales. Deux groupes d'individus de la même espèce, qui doivent donc présenter un vieillissement biologique similaire, pourront avoir une longévité très différente suivant qu'il sont dans la nature ou en captivité, suivant le nombre de prédateurs, etc. Il nous faut donc renoncer à utiliser le simple critère de la longévité pour caractériser le vieillissement biologique.
On pourrait être alors tenté d'utiliser des mesures de prouesses sportives : il est par exemple clair qu'une personne âgée ne peut rivaliser sur un 100 mètres avec un jeune homme. Cependant, le vieillissement n'est pas la seule cause d'une éventuelle baisse de performances : un athlète se mettant à fumer et à ne plus surveiller son alimentation verra sans aucun doute ses résultats baisser spectaculairement, sans que cela soit dû au vieillissement de son organisme. Des problèmes similaires surviennent quand l'on s'intéresse à d'autres fonctions de l'organisme, comme la mémoire ou la fertilité.
En fait, un bon indicateur du vieillissement semble être l'évolution du taux de mortalité, que l'on peut identifier à la probabilité de mourir dans l'année qui vient, à un âge donné. La mortalité suit à l'âge adulte une croissance exponentielle : si elle passe de 1% à 2% en un temps donné, elle passera de 2% à 4% dans le même temps. Bien évidemment, les taux précis de mortalité d'individus d'une même espèce dépendent de nombreux facteurs. Un homme par exemple aura une plus faible probabilité de mourir s'il vit dans une société industrialisée et hautement médicalisée que s'il vit dans une société basée sur l'agriculture et fortement sensible aux épidémies. De même, ces données sont de différentes en temps de guerre ou en temps de paix, ou selon qu'on soit homme ou femme (les femmes présentent dans la quasi-totalité des pays des taux de mortalité inférieurs à ceux des hommes, pour des raisons sans doute biologiques et non sociologiques, mais encore mal comprises). Cependant, le temps de doublement du taux de mortalité semble être une quasi constante à l'intérieur d'une espèce. Chez les hommes, elle est d'à peu près huit ans. Elle peut en fait varier de 7 à 26 ans, mais si l'on ne considère que les hommes de plus de 40 ans et qu'on élimine les morts non dues à des raisons naturelles, on obtient des données qui peuvent encore moins varier.
Ainsi, le temps de doublement du taux de mortalité nous donne un indicateur pertinent pour mesurer la vitesse du vieillissement et comparer les différentes espèces animales : dix jours pour les drosophiles, trois mois pour les souris de laboratoires et huit ans pour les êtres humains. Au sein d'une même espèce, des inégalités face au vieillissement semblent pourtant apparaître : par exemple, il existe des familles dont les membres ont souvent une grande longévité. Bien qu'on ne puisse en être sûr, il ne pourrait s'agir là que d'une meilleure résistance, transmise par les gènes, à certaines maladies, et non le ralentissement du vieillissement. On connaît aussi des maladies, comme le syndrome de Hutchinson-Gilford ou le syndrome de Werner, qui semblent accélérer le vieillissement des personnes qui en sont atteintes : chute de cheveux, plus grand risque de certaines maladies cardio-vasculaires, plus grand risque de cancer pour le syndrome de Werner. Cependant, tous les signes de la sénescence n'apparaissent pas : les malades ne sont pas particulièrement touchés par le diabète ou la maladie d'Alzheimer. Il ne s'agirait donc pas là d'un réel vieillissement accéléré mais de l'apparition de quelques symptômes spectaculaires et tragiques.
Le pourquoi du vieillissement
Le nombre de théories qui s'affrontent dans le domaine du vieillissement est impressionnant : le gérontologue russe Zhores Medvedev en a dénombré plus de 300. Cependant un grand nombre d'entre elles ne s'intéressent pas réellement aux causes, mais plutôt à la mécanique de la sénescence. En ce sens, elles ne sont pas, nécessairement contradictoires et peuvent simplement décrire plusieurs aspects d'un même phénomène.
En matière de théorie du pourquoi du vieillissement, seules trois hypothèses sérieuses se sont affrontées et deux d'entre elles se sont révélées erronées. Cela ne veut malheureusement pas dire que la troisième est la bonne, les chercheurs manquant toujours de données sur la cause du vieillissement.
La théorie du bien de l'espèce
L'évolution d'une espèce, via la sélection naturelle, ne peut survenir que s'il y a un renouvellement des générations. Ce n'est en effet que par la reproduction que des nouvelles combinaisons de gènes apparaissent, fournissant de nouvelles caractéristiques à un individu qui peut ainsi être mieux adapté à son environnement en perpétuel changement. La théorie du bien de l'espèce consiste à dire que le vieillissement permet justement ce renouvellement des générations, les individus les plus âgés mourant pour laisser place aux plus jeunes. C'est donc pour le bien de l'espèce, et non pour le sien propre, qu'un individu vieillit.
Un des faits qui semblent être le plus en faveur de cette théorie est le cas des espèces comme le saumon ou la souris marsupiale Antechinus. Ces animaux se reproduisent une seule fois au cours de leur vie, et subissent aussitôt après un vieillissement accéléré qui entraîne rapidement leur mort (seulement les mâles dans le cas de Antechinus). Il semble naturel d'expliquer cela par la théorie du bien de l'espèce : le vieillissement n'est alors qu'une étape naturelle de la vie, comme la naissance ou la reproduction, cette étape permettant d'accélérer la mort de l'individu et donc de faciliter le renouvellement des générations.
Cependant, cette théorie soulève un problème majeur : elle suppose que l'évolution sélectionne les caractéristiques qui sont favorables au groupe et non à l'individu. Mais cela va à l'encontre des principes de la biologie de l'évolution. Imaginons par exemple une population dans laquelle chaque femelle donne naissance à deux enfants au cours de sa vie, un mâle et une femelle, et que tous les individus meurent de mort naturelle. Cette population est parfaitement équilibrée. La population restant de taille constante, les ressources ne viennent pas à manquer et le renouvellement des générations se fait sans problèmes. Supposons maintenant qu'une mutation survienne et qu'apparaisse chez une femelle un gène qui double la longévité et donc le nombre d'enfants. Ce gène est manifestement mauvais pour le groupe puisque s'il se répand, la population suivra une croissance exponentielle, ce qui posera le problème du manque de ressources. Cependant, ce gène est bon pour l'individu qui vit plus longtemps et transmet à plus d'enfants son patrimoine génétique. Ce que l'évolution prédit, c'est que ce nouveau gène se répande, puisque au fur et à mesure des générations, la part des individus porteur de ce gène sera de plus en plus importante. La sélection se fait donc au niveau de l'individu et non du groupe et la théorie du bien de l'espèce se trouve infirmée.
La théorie du taux de vie
Le métabolisme crée des sous-produits (notamment les radicaux libres) capables d'endommager les cellules et qui sont sans doute un des mécanismes importants du vieillissement. On peut donc penser que la vitesse du métabolisme d'un individu - ce que l'on peut appeler son taux de vie - conditionne son vieillissement. Chaque cellule, de quelque espèce animale qu'elle soit, aurait ainsi le même « capital métabolique », que l'on peut voir comme sa consommation énergétique. Les cellules, et donc l'organisme, vieilliraient ainsi au fur et à mesure de la consommation de ce capital.
Un grand nombre de constatations paraissent soutenir cette théorie : plus un animal est grand, plus il a de cellules, et plus il vit vieux (ce qui, en première approximation signifie qu'il vieillit lentement). Des drosophiles que l'on place dans un environnement plus froid (ce qui réduit d'autant la température interne de ces animaux à sang froid) vivent plus longtemps, et on sait qu'une baisse de température provoque un ralentissement des réactions chimiques qui sont à la base du métabolisme. Enfin, une expérience a montré que si l'on sous-alimentait des rats ou des souris de laboratoire de 30% à 40% par rapport une alimentation non limitée, mais en évitant la malnutrition, ces animaux vivaient 20% à 40% plus vieux. Cela peut s'expliquer par la théorie du taux de vie par un vieillissement ralenti, faute de combustibles au métabolisme.
La conséquence qu'aurait eu cette théorie sur les êtres humains si elle s'était révélée vrai est assez surprenante : il aurait fallu en conclure que pour vieillir plus lentement, et donc vivre plus longtemps, il faudrait manger très peu, mais suffisamment, et éviter tout exercice physique, grand consommateur d'énergie !
Toutefois, un certain nombre d'animaux semblaient faire exception à cette théorie. Les chauves-souris, tout d'abord, proches en taille d'un grand nombre de petits mammifères terrestres, vivent beaucoup plus vieux que ces derniers (alors qu'une souris ne vit que quelques années, une chauve-souris peut vivre plus de vingt ou trente ans, selon son espèce). Cette particularité avait tout d'abord était expliquée par le fait qu'un certain nombre d'espèces de chauves-souris hibernent, et donc ont un métabolisme très ralenti pendant une grande période de leur vie. L'explication ne peut cependant pas tenir, car on s'est aperçu que d'autres espèces de chauves-souris, présentant la même longévité, n'hibernaient pas. Un autre contre-exemple à la théorie du taux de vie vient des oiseaux, qui à taille égale, ont des taux métaboliques et une température corporelle plus élevés que ceux des mammifères. Pourtant, les oiseaux peuvent vivre à un âge très avancé : des corbeaux ou perroquets de plus de 50 ans ne sont pas rares, et il semblerait même que certains perroquets aient dépassé les cent ans. Enfin, on a constaté que les rats sous-alimentés ne diminuaient en fait pas à long terme leur métabolisme. La théorie du taux de vie, toute séduisante aux oisifs qu'elle puisse être, n'est donc pas valable.
La théorie évolutionniste du vieillissement
La maladie de Huntington est une maladie génétique mentale à déclenchement tardif qui conduit à la démence puis à la mort les individus atteints, et dont l'étude a permis au célèbre biologiste Haldane de découvrir un point important pour l'étude des causes du vieillissement. Il a constaté que cette maladie était anormalement répandue, puisqu'elle touche une personne sur 15 000 dans les populations européennes. La sélection naturelle aurait dû quasiment l'éliminer, la ramenant à des taux comme par exemple le syndrome de Hutchinson-Gilford qui touche une personne sur huit millions. En fait, la particularité de cette maladie est qu'au moment où elle frappe les porteurs du gène concerné, ceux-ci ont déjà eu pratiquement tous les enfants qu'ils auraient eu s'ils n'avaient pas été atteints. La maladie n'agissant en général qu'après la période de procréation, la sélection naturelle ne peut pas agir dessus, ce qui explique le nombre élevé de porteurs.
Pour bien comprendre le phénomène, reprenons l'exemple de la population de tubes à essai. Supposons que chaque tube donne naissance à un bébé tube, chaque année de son existence. Même si les tubes ne vieillissent pas, ils seront inévitablement cassés. Par exemple, le taux de casse s'élève à 50% par an. Un tube a alors une chance seize de vivre 4 ans et une chance sur 1024 de vivre dix ans. Supposons enfin qu'apparaisse un gène « suicide » qui provoque l'explosion de son porteur à l'âge de 4 ans. Comme un nombre non négligeable de tubes auront survécu jusqu'à cet âge et qu'ils seront encore en mesure de procréer, ce gène sera sans doute éliminé par la sélection naturelle. Par contre, si l'âge de l'explosion est retardée à 10 ans, son influence sur la population sera quasi-nulle et la sélection naturelle ne favorisera ni défavorisera ce gène. C'est ce qui se passe pour la maladie de Huntington, c'est aussi ce qui pourrait se passer pour le vieillissement : celui-ci n'avait en quelque sorte pas été prévu par l'évolution, ses conséquences n'apparaissant que parce que nous vivons dans un environnement sur-protégé.
Il est aussi possible que les mécanismes (négatifs) associées au vieillissement soient la contrepartie de mécanismes positifs se déroulant plus tôt dans la vie. Ainsi, des expériences sur des cochons d'Inde ont montré que les hormones masculines comme la testostérone pourraient être à l'origine de cancers à un âge avancé. Il pourrait donc y avoir un coût à la reproduction. C'est d'ailleurs à la suite d'une période d'accouplements effrénés que le mâle Antenichus dont nous avons parlé plus haut, meurt brutalement.
Cette théorie évolutionniste du vieillissement a un certain nombre de conséquences : ce sont les animaux subissant une pression environnementale forte qui vieillissent les plus vite. Il serait absurde pour eux de gaspiller de l'énergie à contrer les mécanismes de la sénescence alors qu'ils risquent de mourir rapidement, de la patte d'un prédateur par exemple. Il est donc naturel que les grands animaux vieillissent plus lentement que les petits, que les oiseaux et chauves-souris, disposant de l'atout qu'est le vol, vivent plus longtemps que les mammifères terrestres. Et en effet, on s'aperçoit que les oiseaux qui ne sont pas bon voleurs, comme la volaille, l'autruche ou les pingouins, ont des longévités moins grandes, rapportées à leur taille, que les autres oiseaux (comparer par exemple les longévités de la dinde et du pigeon dans le tableau ci-dessus). Enfin, les saumons meurent après leur reproduction car il valait mieux pour eux consacrer leur énergie à la procréation, plutôt que d'espérer survivre une année de plus. La théorie évolutionniste du vieillissement semble donc expliquer pourquoi la Nature n'a pas plus mis de défenses contre les mécanismes de la sénescence.
Les mécanismes du vieillissement
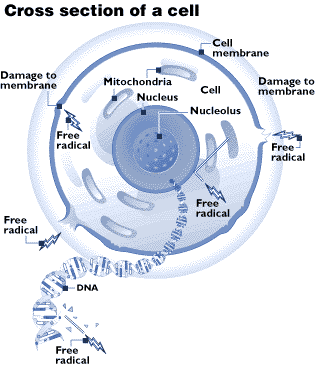
Source :
The Tallahassee
Democrat
L'un des principaux mécanismes connus du vieillissement résulte de la formation, au cours du métabolisme, d'oxydants, dont les radicaux libres. Ces derniers sont des atomes ou molécules qui ont un électron non apparié sur leur couche externe et qui réagissent donc très fortement avec d'autres molécules, pour se stabiliser et former de nouveaux radicaux libres. Ce processus d'oxydation peut endommager n'importe quelle partie de la cellule, comme les mitochondries qui sont le siège du métabolisme ou l'ADN, siège de l'information génétique. Des mutations peuvent ainsi être provoquées, et on sait que ce sont de telles mutations qui sont à l'origine de la formation de cellules immortelles, c'est-à-dire un cancer. Face à cette menace, l'organisme produit une quantité importante d'antioxydants censés stabiliser les radicaux libres. On sait que les vitamines A, C et E sont de bons antioxydants, mais leur intérêt in vivo dans la lutte contre le vieillissement n'est pas avéré. L'organisme se défend également en réparant les séquences d'ADN endommagées (cela est possible car les brins d'ADN viennent par paire, la séquence d'un brin se déduisant de la séquence de l'autre). Cependant, si la réparation n'a pas le temps de se faire avant un dédoublemement de la cellule (une mitose), la mutation devient irrémédiable.
Un autre processus mis en jeu par le vieillissement est la réaction de glycosylation découverte par le chimiste français Louis Maillard : les molécules de glucose présentes dans l'organisme peuvent se combiner à différentes protéines, modifiant ainsi leur structure. Quand on sait que les fonctions des protéines dépendent justement le plus part du temps de leur structure, on comprend les dommages qui peuvent être occasionnés. C'est, semblerait-il, ce mécanisme qui serait en jeu dans la maladie d'Alzheimer. Là encore, le corps a prévu des défenses : certains phagocytes semblent spécialisés dans la destruction des protéines endommagées. Il est à noter que les deux mécanismes précédemment cités agissent de concert, les radicaux libres accélérant la glycosylation et cette dernière pouvant engendrer des radicaux libres.
Conclusion - Perspectives
L'étude du vieillissement est encore un domaine scientifique jeune et prometteur. Si un grand nombre d'hypothèses ont été émises, on manque d'une théorie causale claire. Quant à d'éventuelles applications médicales dans le ralentissement du vieillissement, nous n'en sommes pas encore là. Les produits miracles qui promettent des effets spectaculaires dans cette direction ont autant de validité scientifique que la fontaine de jouvence. Cependant, tous les espoirs sont permis dans ce secteur en pleine ébullition. Que ce soit souhaitable éthiquement ou non, l'homme de 150 ans n'est certes pas pour demain, mais peut-être pour après-demain...
Bibliographie
- Austad, Steven N. ; Why we age ; New York ; John Wiley & Sons, Inc. ; 1997.
- « Dossier » ; La Recherche ; juillet-août 1999 ; n°322.
Merci à Jacques Tréton pour ses conseils bibliographiques et à Édouard Maurel-Segala pour sa relecture.